
◆ 潜水型(せんすいがた)の鳥の進化 ◆
潜水時に翼(つばさ)を使う鳥は、3つの独立(どくりつ)した進化系から発生している。
図Ⅰの横の系統樹(けいとうじゅ)のように、南半球では、ウミツバメ類からモグリウミツバメ類が、ペンギン類はミズナギドリの祖先(そせん)から派生(はせい)したと考えられる。
北半球では、ウミスズメとその近縁種(きんえんしゅ)の中で知られており、現存、絶滅種(ぜつめつしゅ)のどちらもシギ・チドリなどの祖先から派生している。
北半球において、ペンギンに匹敵(ひってき)する鳥は19世紀に絶滅した「オオウミガラス」を含むウミガラスの仲間である。
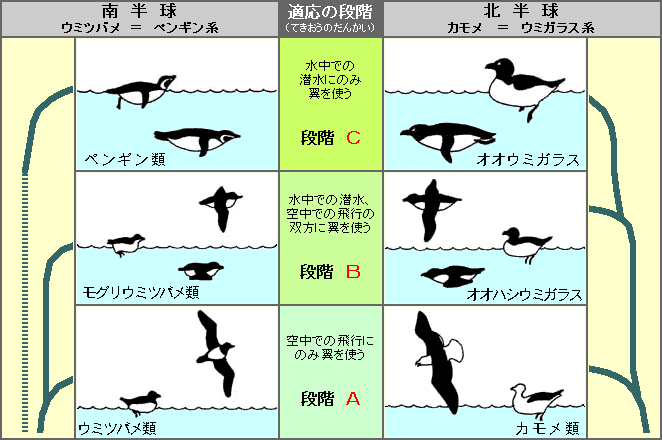
▲図Ⅰ 翼で推進(すいしん)する潜水性の鳥の二原型の進化における平行的な適応の段階。
左は南半球の原型、右は北半球の原型 (Storer, 1960 より)
1960年にロバート・ストラーは、翼で推進する潜水様式(せんすいようしき)の鳥類が、北半球と南半球でそれぞれの進化の段階で生態学的(せいたいがくてき)に同等(どうとう)である、と論じている。(図Ⅰ)
生物の生態系における位置を「生態的地位(ニッチ)」と呼ぶ。
どのような生活条件の元で、どのような食生活を営んでいるかということである。
同じ種類ではないのに、南北両半球で、同じニッチを占める海鳥が、同じような姿、生活形態を示すように進化した。(平行進化(へいこうしんか))
最初にあらわれた海鳥は、翼を空を飛ぶ場合にのみ使う海鳥。(段階A)
北半球ではカモメ類が、南半球ではウミツバメ類が現れた。
その次に水中と空中の両方で翼を使う海鳥の出現。(段階B)
そして現段階での最終段階(さいしゅうだんかい)として、潜水時にのみ翼を使う海鳥が現れる。(段階C)
北半球では絶滅したオオウミガラス、南半球ではご存知のペンギン類である。
南北両半球で、それぞれに関わりがあったわけではないのに、同じようなニッチを占(し)める海鳥がそれぞれに同種の海鳥のような進化段階を示している。
同じような平行進化の例は、他の動物でも良く見られる。
面白い例としては、オーストラリアの有袋類(ゆうたいるい)である。
ご存知のように、カンガルーやコアラに代表される有袋類はオーストラリアにのみ見られ、長い間他の陸地から隔絶(かくぜつ)されたために、独自(どくじ)の進化を示してきた結果である。
その有袋類と、他の大陸の動物で、同じようなニッチを占めている動物同志で、ほとんど同種と見まがうようなものが多い。
フクロモモンガ=モモンガ、フクロリス=リス、フクロウサギ=ウサギ、フクロトビネズミ=トビネズミ、フクロオオカミ=オオカミ、、等などである。(それぞれ前者が有袋類)
突然変異(とつぜんへんい)が進化の原動力(げんどうりょく)だと言うのなら、なぜこのような平行進化の例が頻繁(ひんぱん)に見られるのかが疑問である。
同じくストラーは、ペンギン類の翼を生み出したと思われる進化の過程(かてい)を、異なる鳥類グループから5例を引用(いんよう)して説明している。(図Ⅱ)
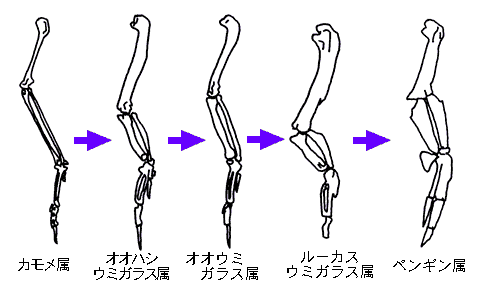
▲図Ⅱ ペンギン類の翼が進化するまでに経た段階と類似(るいじ)の翼の適応段階(てきおうだんかい)
(なお図Ⅱは、あくまで適応の段階を比較したものであって、
それぞれの鳥類から順次(じゅんじ)進化していったわけではないので注意。
ペンギンの祖先がルーカスウミガラスなのではない。 )